5
Objectifs
- Expliquer l’importance du noyau dans une cellule ;
- Identifier et décrire dans l’ordre chronologique les différentes phases de la mitose ;
- Expliquer le comportement des chromosomes ;
- Représenter et nommer les différentes parties d’un chromosome ;
- Établir une relation entre la variation du taux d’ADN et le comportement des chromosomes ;
- Décrire et schématiser la structure de l’ADN ;
- Définir et expliquer les mécanismes de réplication, transcription et traduction ;
- Expliquer que la mitose assure la croissance des êtres vivants et la pérennité des espèces ;
- Énoncer la théorie cellulaire.
INTRODUCTION
Une cellule vivante doit se reproduire, c’est-à-dire engendrer d’autres cellules identiques à la cellule initiale. Ceci suppose l’existence dans la cellule des molécules capables de s’autoreproduire autour desquelles toute la structure de la cellule s’organise.
Des observations microscopiques et des expériences permettent de déterminer les mécanismes cellulaire et moléculaire de la division cellulaire.
1- L’importance du noyau dans la vie cellulaire
Expériences et résultats : L’expérience de mérotonie (section de la cellule) chez l’amibe montre que le morceau nucléé survit et régénère le fragment amputé tandis que le morceau anucléé meurt.
En 1939, FONTBRUNE arrive à extraire le noyau d’une amibe : le noyau isolé et l’amibe énucléée meurent. Il greffe un nouveau noyau provenant d’une amibe de la même espèce sur l’amibe énucléée : il constate que l’amibe recombinée retrouve toute son activité vitale.
Interprétation : L’expérience de mérotonie montre que les fonctions vitales ne s’observent qu’en présence du noyau. L’expérience de greffe confirme le rôle vital du noyau mais précise qu’une cellule ne peut pas vivre avec son seul noyau.
Conclusion : Le noyau est nécessaire à la vie de la cellule.
2- La division cellulaire (mitose)
L’observation au microscope des cellules de la pointe de racine ou de la moelle rouge des os montre que la région du noyau présente une grande variété : elle est soit compacte et granuleuse ou au contraire semble disparaître pour être remplacée par des structures en forme de filaments (document 4.1).
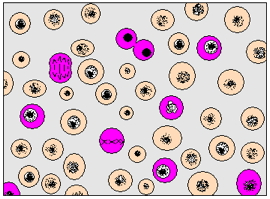
La présence des cellules sœurs séparées par une mince paroi ou reliées par un fin pont cytoplasmique suggère que dans ces zones, les cellules se divisent et on peut reconstituer les différentes phases de ce phénomène.
2.1- Les étapes de la mitose chez une cellule animale (document 4.2)
La mitose est un phénomène continu mais on peut distinguer quatre phases aux limites peu arbitraires :
a- La prophase (durée : 15 à 60 minutes)
Elle se caractérise par :
- L’organisation de la chromatine en filaments de plus en plus individualisés, de plus en plus courts, épais et colorables, les chromosomes. Chaque chromosome apparaît fissuré (clivé) longitudinalement en deux brins ou chromatides reliés par un point non colorable appelé centromère;
- La disparition de l’enveloppe nucléaire et du (des) nucléole(s);
- Le dédoublement de la paire de centrioles formant le centrosome et chaque paire s’éloigne l’une de l’autre en direction des pôles ; les microtubules édifiés autour de chaque centrosome élaborent une figure particulière appelée aster;
- La formation du fuseau achromatique (fuseau de division) qui est un appareil résultant de l’organisation des microtubules en fibres dirigées d’un pôle vers l’autre (fibres polaires) ou disposées perpendiculairement aux chromosomes (fibres chromosomiques) ou disposées autour des centrioles (fibres astériennes).
b- La métaphase (durée : quelques minutes)
Les chromosomes sont de plus en plus condensés et individualisés : on peut réaliser le caryotype de la cellule.
Les centromères se regroupent dans le plan équatorial du fuseau de division. L’ensemble de chromosomes clivés ainsi rangés dans le plan médian de la cellule forme une figure appelée plaque équatoriale.
c- L’anaphase (durée : 2 à 10 minutes)
Les deux chromatides de chaque chromosome métaphasique s’écartent l’une de l’autre après coupure de chaque centromère.
Deux lots identiques de chromosomes à une chromatide migrent chacun vers l’un des pôles du fuseau de division. Cette migration due à la rétraction des fibres chromosomiques est appelée ascension polaire.
d- La télophase (durée : 15 à 60 minutes)
Un noyau se forme au niveau de chaque pôle par désindividualisation des chromatides qui retournent à l’état de chromatine diffuse, reconstitution de l’enveloppe nucléaire et du (des) nucléole(s).
Le fuseau de division disparaît. Le cytoplasme se divise entre les deux noyaux fils par étranglement de la membrane cytoplasmique : c’est la cytodiérèse.
Remarque : particularités de la mitose chez les cellules végétales.
La mitose d’une cellule végétale se déroule dans les grandes lignes comme celle d’une cellule animale à deux différences près :
- L’absence des asters qui sont remplacés par des zones plus claires du cytoplasme appelées calottes polaires autour desquelles s’organise le fuseau de division ;
- La cloison cellulaire qui sépare les deux cellules filles issues de la division se forme à partir des vésicules golgiennes qui s’alignent dans le plan médian et fusionnent pour donner une plaque appelée phragmoplaste, ébauche de la paroi pectocellulosique.
Conclusion : La mitose est un phénomène universel qui aboutit à la formation de deux cellules filles identiques entre elles et identiques à la cellule de départ.
Comment expliquer le maintien de la constance du nombre chromosomes d’une cellule à l’autre au cours de la mitose ?
2.2- La structure d’un chromosome (document 4.3)
Activité : observation des documents (pages 30 et 31, SVT 1ère S, Ed. Hatier)
- Annoter et donner un titre à chaque figure.
- Identifier les constituants chimiques du chromosome et de la chromatine.
- Expliquer le mode d’agencement de ces constituants.
- Expliquer le comportement des chromosomes observé au cours de la vie cellulaire (visibles en mitose et invisibles en interphase c’est-à-dire entre deux mitoses).
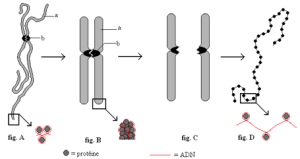
Chaque chromosome est constitué de deux composants majeurs :
- L’acide désoxyribonucléique (ADN) ;
- Les protéines (histones et non histones).
La molécule d’ADN filiforme s’enroule autour des protéines histones, formant un nucléofilament ou chaîne nucléosomique (collier de perles). Selon le degré de spiralisation du nucléofilament, le chromosome est plus ou moins visible. Les protéines jouent un rôle essentiel dans ce comportement.
La spiralisation est maximale à la métaphase : les chromosomes sont alors nettement individualisés.
Conclusion : le chromosome est une structure permanente du noyau cellulaire plus ou moins visible selon le degré de spiralisation du nucléofilament.
2.3- L’évolution du taux d’ADN au cours de la mitose (Exercice 1)
Les cellules qui se divisent régulièrement effectuent un cycle appelé cycle cellulaire que l’on peut diviser en deux phases : mitose et interphase qui alternent régulièrement.
Le dosage de la quantité d’ADN par noyau cellulaire au cours d’un cycle cellulaire permet de dresser le tableau suivant :
| Temps | 0 | 1 h | 1 h 45 | 1 h 50 | 3 h | 5 h 30 | 7 h | 9 h | 10 h | 12 h | 13 h 45 | 13 h 50 | 15 h |
| Quantité d’ADN (unités arbitraires) | 8 | 8 | 8 | 4 | 4 | 4 | 5 | 7 | 8 | 8 | 8 | 4 | 4 |
- Tracer la courbe montrant l’évolution du taux d’ADN au cours du temps ;
- Indiquer le début et la fin d’une mitose, ainsi que les différentes phases de la mitose, sachant que pour ces cellules, la mitose dure une heure environ et que la prophase et la métaphase représentent 75 % du temps de la division ;
- Evaluer la durée d’un cycle cellulaire ;
- Interpréter la courbe obtenue ;
- Schématiser sur chaque palier horizontal le comportement d’un chromosome.
La quantité d’ADN dans le noyau varie avec le temps. Elle double entre deux divisions consécutives (interphase). Cette phase de synthèse de l’ADN appelée phase S (synthesis) correspond à la fabrication d’une deuxième chromatide par chaque chromosome.
La quantité d’ADN est réduite de moitié au cours de la mitose. Cette réduction s’explique par le fait que l’anaphase correspond à la division chaque chromosome à deux chromatides en deux chromosomes à une chromatide chacun.
Un cycle cellulaire peut être subdivisé en quatre phases :
- Phase G1 (growth) qui correspond à une première phase de croissance de la cellule ;
- Phase S au cours de laquelle la cellule double son stock d’ADN ;
- Phase G2 pendant laquelle la croissance de la cellule se poursuit.
- Phase M qui correspond à la mitose au cours de laquelle les deux copies d’ADN se repartissent équitablement entre les deux cellules filles.
3- Structure des acides nucléiques (ADN et ARN)
3.1- Structure de l’ADN (document 4.4)
Activité : exploitation des résultats d’expériences.
Voici un fragment d’une chaîne (ou brin) d’une molécule d’ADN : c’est un polynucléotide.
- Identifier les différents constituants de la molécule d’ADN ;
- Décrire le mode d’assemblage de ces différents constituants et identifier les motifs qui se répètent ;
- Citer alors les différents types de nucléotides et justifier le terme polynucléotide attribué à l’ADN.
|
Ø Quelles constatations faites-vous sur la composition en bases azotées de l’ADN des différents organismes étudiés ? Ø Dans une molécule d’ADN, quelles sont les bases complémentaires ? Schématiser la chaîne nucléotidique complémentaire à celle proposée. Ø Décrire la structure de la molécule d’ADN ainsi obtenue. Ø Compléter la représentation simplifiée du fragment de la molécule d’ADN proposé. |
Le tableau suivant donne la quantité relative des quatre bases dans les échantillons d’ADN des cellules de différents organismes.
| Organismes | Pourcentage de chaque base dans l’ADN | |||
| A | G | C | T | |
|
Homo sapiens Mouton Poule Saumon Criquet Escherichia coli Phage T2 |
30,9 29,3 28,8 29,7 29,3 24,7 32,5 |
19,9 21,4 20,5 20,8 20,5 26,0 17,9 |
19,8 21,0 21,5 20,4 20,7 25,7 17,0 |
29,4 28,3 29,3 29,1 29,3 23,6 32,6 |
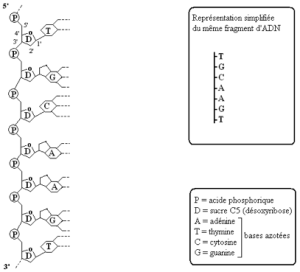
L’ADN est constitué d’un grand nombre d’unités élémentaires appelés nucléotides.
Chaque nucléotide est formé par :
- Une molécule d’acide phosphorique (H3PO4) ;
- Une molécule de sucre à 5 atomes de carbone (pentose) : le désoxyribose;
- Une molécule de base azotée : adénine, thymine, guanine ou cytosine.
L’acide est relié au cinquième atome de carbone du sucre et la base est fixée sur le premier atome de carbone (structure primaire de l’ADN).
Il existe donc quatre types de nucléotides : thymidine, cytidine, guanosine et adénosine.
Les nucléotides sont unis les uns aux autres par les molécules d’acides phosphoriques qui établissent des liaisons covalentes avec le troisième atome de carbone du sucre du nucléotide suivant (structure secondaire de l’ADN) : on obtient ainsi une chaîne polynucléotidique pouvant compter plusieurs millions de nucléotides.
L’originalité de la molécule d’ADN tient à sa structure tertiaire : elle est constituée par deux chaînes de nucléotides qui se font face et se stabilisent par des liaisons hydrogènes établies entre leurs bases. Cette structure à deux chaînes ou brins est appelée structure bicaténaire.
Les deux chaînes décrivent une spirale dans l’espace, l’une autour de l’autre : c’est la structure à double hélice (Watson et Crick, 1953).
Les deux brins sont unis par les bases qui se font face et qui sont complémentaires l’une de l’autre :
- A est toujours relié à T par deux liaisons hydrogènes ;
- C est toujours relié à G par trois liaisons hydrogènes.
Conséquence, la relation liant les quantités des différentes bases azotées de l’ADN est la suivante :
![]()
3.2- Structure de l’ARN (document 4.5)
Activité :
Voici une portion de la molécule d’ARN.
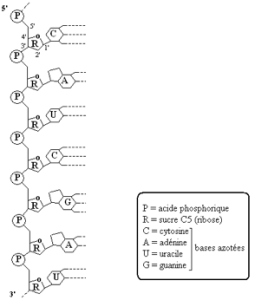
|
– Comparer les molécules d’ADN et d’ARN sous la forme d’un tableau. – Donner une représentation simplifiée du segment d’ARN proposé. |
Le tableau suivant donne la quantité relative des quatre bases azotées dans un échantillon d’ARN des cellules de différents organismes.
| Organisme | Pourcentages de chaque base dans l’ARN | |||
| A | G | C | U | |
|
Homme Triton Truite VIH |
31,9 24,7 25,5 33,7 |
20,1 25,2 23,5 17,8 |
21,6 16,3 18,9 23,5 |
26,4 33,8 32,1 25,0 |
Comme l’ADN, l’ARN est un polynucléotide. Les différences entre les deux molécules sont les suivantes :
- – La molécule de sucre de chaque nucléotide de l’ARN est le ribose (C5H10O5) ;
- – L’uracile remplace la thymine dans l’ARN ;
- – La molécule d’ARN est constituée d’un seul brin de nucléotides : sa structure est dite monocaténaire.
- – D’autres expériences montrent que l’ADN se localise dans le noyau essentiellement alors que l’ARN se trouve dans le noyau et le cytoplasme.
Remarque : l’ADN se trouve dans les chloroplastes et les mitochondries où elle présente une forme circulaire.
4- Le rôle des acides nucléiques
La conservation du nombre de chromosomes et la transmission fidèle de l’information génétique d’une cellule à l’autre au cours de la mitose reposent sur les propriétés de la molécule d’ADN qui peut s’autoreproduire et porter un message codé.
4.1- La duplication ou réplication de l’ADN
La structure de l’ADN rend compte de sa capacité à se répliquer.
a- La synthèse de l’ADN : où et quand ? (exercice 2)
Activité :
La synthèse de l’ADN peut être mise en évidence par autoradiographie. Les molécules radioactives sont révélées par la présence des grains d’argent.
La thymidine tritiée (3H) est injectée à un papillon. L’autoradiographie des tissus est ensuite réalisée. Certaines cellules contiennent des grains d’argent, d’autres non. Les grains d’argent se localisent essentiellement dans le noyau.
Interpréter les résultats obtenus : qu’indique la présence des grains d’argent ? Comment expliquer que parmi les cellules prélevées, seules certaines présentent les grains d’argent ?
A partir des connaissances acquises sur la composition de la molécule d’ADN, citer tous les acteurs susceptibles d’intervenir dans sa synthèse.
La synthèse d’ADN se fait dans le noyau et à des périodes bien précises de la vie cellulaire (phase S de l’interphase).
Cette synthèse nécessite des nucléotides libres, de l’énergie (ATP) et des enzymes.
b- Mécanisme de la réplication de l’ADN
Deux hypothèses sont à éprouver :
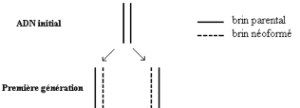
Première hypothèse : l’hypothèse d’une réplication conservative de l’ADN.
Une molécule d’ADN sert intégralement de modèle à la molécule néoformée.
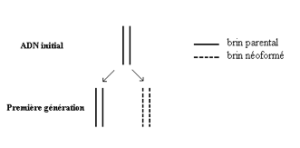
Deuxième hypothèse : hypothèse d’une réplication semi-conservative.
Les deux brins de la molécule mère se séparent et chacun sert de matrice à la formation du brin complémentaire.
Expérience de Meselson et Stahl.
Principe : La destinée des molécules d’une cellule peut être suivie en utilisant des isotopes marqueurs, par exemple 15N, l’isotope lourd de l’azote 14N. L’azote 15N s’incorpore sans discrimination dans tous les composés azotés de la cellule, y compris dans l’ADN. Les molécules d’ADN marquées avec 15N seront plus lourdes que celles contenant 14N, elles pourront ainsi être séparées par centrifugation en gradient de densité et leur densité pourra être évaluée.
Protocole : Des bactéries sont cultivées pendant plusieurs générations sur un milieu contenant des sels d’ammonium tous marqués par 15N. Quand tout l’ADN des bactéries contient du 15N, on transfère ces dernières sur un milieu à 14N. Elles poursuivent leur division de façon synchrone ; on prélève des bactéries à des temps correspondant au dédoublement de la population et on mesure la densité de l’ADN extrait de ces bactéries.
Résultats : (document 4.6)
Résultats prévisibles après une génération sur milieu 14N.
| Hypothèse 1 | Hypothèse 2 | |
| Densité de l’ADN de première génération |
50 % ADN lourd 50 % ADN léger |
100 % ADN « hybride » |
Résultats obtenus par Meselson et Stahl

- Dans le cadre de chaque hypothèse, prévoir les résultats susceptibles d’être obtenus après deux générations sur milieu 14
- Sachant que les bactéries n’ont qu’un seul chromosome, donc une seule molécule d’ADN par cellule, représenter les molécules d’ADN de la deuxième génération (utiliser les figures proposées dans les hypothèses : _______ brin parental et ———- brin néoformé).
- Les informations tirées des résultats obtenus vous permettent-elles de réfuter une des deux hypothèses ? Justifier.
- En tenant compte de la complémentarité des bases azotées de l’ADN, représentez dans le cadre de l’hypothèse retenue, la réplication d’une molécule d’ADN en utilisant la représentation simplifiée du paragraphe 3.1 (structure de l’ADN).
- Comparer l’information codée portée par la molécule d’ADN initiale et les molécules filles.
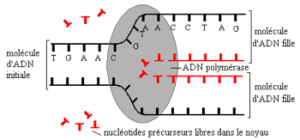
La structure de l’ADN à deux brins reliés par des liaisons de faible énergie et l’existence de bases complémentaires ont permis d’imaginer un mécanisme de doublement des molécules d’ADN par réplication semi-conservative (document 4.7).
Les expériences de Meselson et Stahl montrent que les molécules d’ADN filles sont en effet toutes formées d’un brin parental et d’un brin complémentaire néoformé.
Lors de la réplication semi-conservative, chaque molécule d’ADN s’ouvre en des points variés par rupture des liaisons hydrogènes (yeux de réplication). Des nucléotides complémentaires viennent se placer en face des nucléotides des deux brins de la molécule ouverte. Des liaisons covalentes créées entre les différents nucléotides assurent la formation des nouveaux brins. Ce processus nécessite de l’énergie et un complexe enzymatique : l’ADN polymérase.
Les deux molécules filles hybrides ont une séquence nucléotidique identique entre elles et identiques à la molécule parentale. Ainsi est conservée l’information génétique au cours de la réplication.
4.2- La transcription de l’ADN
De nombreux faits et expériences ont montré que l’intermédiaire entre l’information génétique contenue dans le noyau et le lieu de synthèse des protéines était une molécule d’ARNm.
a- Localisation de l’ARN (document 4.8)
Des cellules de Tetrahymena (protozoaire) sont cultivées en présence de l’uracile radioactif pendant quelques minutes, puis transférées sur un milieu non radioactif. Les figures suivantes représentent des autoradiogrammes de cellules fixées au moment du changement de milieu (figure 1) et quelques heures après (figure 2).
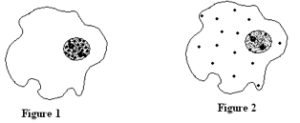
A l’aide des informations tirées de ces figures, montrer que l’ARN présente des caractéristiques nécessaires au rôle qui lui est attribué.
Les différences entre les deux figures s’expliquent par le fait que l’incorporation de l’uracile dans les molécules d’ARN s’effectue dans le noyau ; les molécules d’ARN formées passent ensuite dans le cytoplasme.
L’ARN peut ainsi porter le message génétique du noyau au cytoplasme d’où le nom ARN messager (ARNm).
b- Mécanisme de la transcription (documents 4.9 et 4.10)
L’insuline est une hormone protéique synthétisée par les cellules β du pancréas endocrine. Elle est formée de deux chaînes A et B comportant respectivement 21 et 30 acides aminés.
Voici la séquence des nucléotides du fragment d’ADN portant l’information génétique pour la synthèse de la chaîne B de l’insuline (gène).
Brin 1 : TTT GTG AAC CAA CAC CTG TGC GGC TCA CAC CTG GTG GAA GCT CTC TAC CTA GTG TGC GGG GAA CGA GGC TTC TTC TAC ACA CCC AAG ACC
Brin 2 : AAA CAC TTG GTT GTG GAC ACG CCG AGT GTG GAC CAC CTT CGA GAG ATG GAT CAC ACG CCC CTT GCT CCG AAG AAG ATG TGT GGG TTC TGG
La séquence en nucléotides de l’ARNm extrait de cellules β des îlots de Langerhans pendant une période de synthèse d’insuline est la suivante (NB : on n’a représenté que le fragment d’ARNm codant pour la chaîne B).
UUU GUG AAC CAA CAC CUG UGC GGC UCA CAC CUG GUG GAA GCU CUC UAC CUA GUG UGC GGG GAA CGA GGC UUC UUC UAC ACA CCC AAG ACC
L’analyse des séquences nucléotidiques proposées permet-elle de confirmer le rôle de l’ARNm ? Justifier.
En transposant vos acquis sur les mécanismes de synthèse de l’ADN, identifier le brin d’ADN qui a servi de matrice et proposer un modèle qui explique la formation d’un ARNm.
L’ARNm a une séquence nucléotidique complémentaire de l’un des deux brins de l’ADN. Ce brin est appelé brin transcrit ou brin non codant.
On en déduit que l’ARNm est une copie de l’un des brins de l’ADN.
Si on bloque la formation de cet ARN par des substances chimiques, la synthèse des protéines cesse. C’est donc l’ARN qui copie l’information portée par l’ADN et transporte ce message jusqu’au lieu de synthèse des protéines.
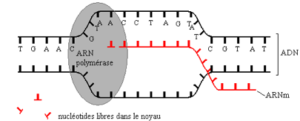
La transcription est l’opération qui consiste à copier une séquence nucléotidique de l’ADN en ARNm. Elle a lieu essentiellement pendant les phases G1 et G2 de l’interphase. Elle nécessite des nucléotides libres, de l’énergie et un complexe enzymatique : l’ARN polymérase. Il faut aussi des signaux spécifiques (séquences nucléotidiques particulières) pour indiquer sur l’immense molécule d’ADN, les début et fin de la portion à transcrire (gène).
Les principales étapes de la transcription sont les suivantes (document 4.10):
- – Ouverture de la molécule d’ADN au niveau de la séquence nucléotidique à copier en ARN ;
- – Mise en place des nucléotidiques d’ARN face à des nucléotides d’ADN d’un seul brin qui sert de matrice par le jeu de la complémentarité des bases :
| ADN | ARN |
| A T C G |
U A G C |
- Liaison entre les ribonucléotides et formation d’un brin d’ARN.
L’ARNm formé se détache rapidement du brin d’ADN transcrit et migre vers le cytoplasme. Les deux brins se réapparient.
4.3- Le code génétique
a- Expériences de Nirenberg et Ochoa
Des extraits obtenus à partir des bactéries sont mis en présence des acides aminés radioactifs ; leur incorporation éventuelle peut ainsi être suivie. A cette préparation, est ajouté un ARNm de synthèse formé entièrement des nucléotides d’uridine.
Ce polymère (poly U) détermine in vitro la synthèse d’un polypeptide formé d’un seul acide aminé : la phénylalanine.
Il semble donc que les nucléotides à uracile correspondent à la phénylalanine.
D’autres expériences montrent que les ARNm poly C provoquent la synthèse des polypeptides formés uniquement de proline ; un ARNm poly A détermine l’assemblage des molécules de lysine.
On peut en conclure qu’il existe un système de correspondance entre la séquence des nucléotides de l’ARNm et la séquence des acides aminés de la protéine : c’est le code génétique.
b- Le code génétique
Sachant que l’ADN ou l’ARN contient quatre types de nucléotides et que la protéine est formée par 20 acides aminés différents, le problème est de savoir comment à partir d’un alphabet de quatre lettres, nous pouvons composer 20 mots différents pour désigner les 20 objets que sont les acides aminés.
Hypothèse 1 : un nucléotide désigne un acide aminé. On peut désigner quatre acides aminés ; ce qui est insuffisant.
Hypothèse 2 : deux nucléotides désignent un acide aminé. On peut désigner 42 = 16 acides aminés différents ; ce qui reste insuffisant.
Hypothèse 3 : trois nucléotides désignent un acide aminé. On peut former 43 = 64 combinaisons différentes, soit plus de mots qu’il n’en faut pour désigner les 20 acides aminés.
De nombreuses expériences montrent que la troisième hypothèse est la bonne :
- – L’unité de l’information génétique est un triplet de nucléotides correspondant à un codon ;
- – Plusieurs codons désignent un seul et même acide aminé : ce sont des codons synonymes;
- – Trois codons ne désignent aucun acide aminé (UAA, UAG et UGA) : ce sont des codons non-sens, ou codons stop ou codons de ponctuation. Ce sont des signaux d’arrêt de la traduction (document 4.11).
c- Les caractères du code génétique
- – Le code génétique est redondant ou dégénéré: plusieurs codons désignent un même acide aminé ;
- – Le code génétique est universel à quelques exceptions près : la signification d’un codon est la même chez presque tous les êtres vivants ;
- – Le code génétique est non chevauchant: un nucléotide n’est impliqué que dans un seul codon.
4.4- La traduction
C’est la synthèse des protéines c’est-à-dire l’expression de l’information portée par l’ARNm en une protéine.
a- Conditions
Pour que la traduction ait lieu, il faut que :
- – De l’ARNm ait été fabriqué ;
- – Tous les acides aminés nécessaires soient présents dans la cellule ;
- – De l’énergie soit fournie (ATP) ;
- – Tous les outils soient présents : des ARNt (ARN de transfert), des enzymes permettant de charger un acide aminé sur son ARNt, des ribosomes.
Ces outils sont universels c’est-à-dire qu’ils peuvent servir à la fabrication de n’importe quelle protéine, étrangère à la cellule ou non.
b- Rôle des ARNt
Ils jouent le rôle d’adaptateurs et déterminent la mise en place des acides aminés dans la chaîne polypeptidique grâce à la complémentarité de leur triplet de nucléotides (anticodon) avec un codon de l’ARNm.
Seule cette complémentarité intervient quel que soit l’acide aminé chargé par l’ARNt. Le chargement de l’acide aminé sur son ARNt nécessite une enzyme (l’amino acyl ARNt synthétase) et de l’énergie (ATP) (document 4.12).
c- Rôle des ribosomes
Ils ont une organisation les rendant aptes à se fixer sur un ARNm. Ils assurent le positionnement correct des ARNt chargés vis-à-vis des codons de l’ARNm.
Ils possèdent les enzymes qui catalysent la liaison peptidique entre les acides aminés.
d- Mécanisme de la traduction
La synthèse d’une chaîne polypeptidique comporte des étapes successives suivantes : l’initiation, l’élongation et la terminaison.
i- L’initiation (document 4.13)
La synthèse débute par la mise en place au niveau du codon AUG de l’ARNm de la petite sous-unité ribosomiale, d’un ARNt porteur d’un premier acide aminé (la méthionine), puis de la grosse sous-unité ribosomiale.
ii- L’élongation (document 4.14)
Elle s’effectue par l’adjonction de nouveaux acides aminés. Cet accrochage successif se fait en trois temps :
- – Association codon-anticodon ;
- – Transfert d’énergie permettant la formation d’une liaison peptidique entre deux acides aminés ;
- – Déplacement relatif du ribosome et de l’ARNm sur une longueur d’un codon, dans le sens 5’ → 3’ : on parle d’une lecture unidirectionnelle et orientée.
iii- La terminaison (document 4.15)
C’est l’arrêt de la biosynthèse qui survient lors de la lecture d’un codon de ponctuation. Elle entraîne la dissociation du complexe ARNm – sous-unités ribosomiales – chaîne polypeptidique néoformée.
Remarque : la molécule d’ARNm comprend en réalité des séquences nucléotidiques codantes traduites en acides aminés (exons) précédées en amont et suivies en aval par des séquences nucléotidiques non codantes (introns). Ces dernières assurent la stabilité des ARNm et la lecture correcte du message par les ribosomes.
5- Devenir des protéines synthétisées : rôle du REG et du dictyosome
Les protéines synthétisées ont une double destination :
- elles peuvent être intégrées dans la cellule qui les a fabriqué pour être utilisées par cette dernière (cas des protéines membranaires);
- elles peuvent être exportées hors de cette cellule et être utilisées ailleurs (cas des hormones, des enzymes digestives).
Le mécanisme de triage est assez complexe.
5.1- Un véritable « code postal » en fonction de la destination.
a- L’élongation de nombreuses chaînes polypeptidiques débute par la synthèse d’une séquence de 15 à 30 acides aminés appelée peptide signal qui dirige la protéine vers tel organite ou tel autre. Ainsi :
- les protéines destinées à la mitochondrie ou au chloroplaste ont un peptide signal reconnu par les récepteurs spécifiques portés par la membrane de ces organites ;
- les protéines destinées à l’exportation ont un peptide signal qui déclenche la fixation du ribosome, lecteur de l’ARNm, sur la membrane du RE et la protéine synthétisée rentre directement dans la lumière du RE.
Dans tous les cas, le peptide signal est excisé (amputé) par une enzyme spécifique dès que la protéine a atteint l’organite auquel elle est destinée.
b- Si une protéine n’a pas de peptide signal, elle doit rester dans le hyaloplasme (cytoplasme – organites). C’est le cas de certaines enzymes et protéines de structure.
5.2. Mécanisme d’exportation des protéines (document 4.16)
Les protéines destinées à être exportées sont introduites directement dans le RE au moment même de leur synthèse :
- le RE ‘’bourgeonne’’ des vésicules contenant les polypeptides en transit ;
- ces vésicules s’assemblent pour former les saccules golgiens où les chaînes polypeptidiques subissent des remaniements complexes : coupures de fragments, fixation des molécules comme les glucides (cas des glycoprotéines). Les protéines achevées sont ‘’emballées’’ dans des vésicules golgiennes qui vont s’accumuler pour former des volumineuses vésicules de sécrétion ;
- les vésicules de sécrétion fusionnent avec la membrane cytoplasmique de la cellule pour déverser leur contenu hors de la cellule. Ce mécanisme de sécrétion est appelé exocytose.
Remarque : l’exocytose a pour conséquence directe l’augmentation de la surface de la membrane plasmique. Cette création permanente de la membrane plasmique est compensée par des mécanismes inverses (dits d’endocytose) pendant lesquels la membrane plasmique bourgeonne pour faire entrer dans la cellule des substances d’origine externe. La membrane plasmique est ainsi soumise à un renouvellement rapide et permanent.
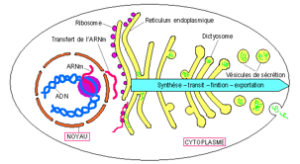
De l’ADN à la protéine
TRAVAIL DEMANDE : Les biologistes disent que les dictyosomes se forment ‘’à partir’’ du REG. Après avoir examiné attentivement cette figure, émettre une hypothèse pour expliquer comment cela est possible.
Résumer en quelques phrases les grandes étapes permettant de passer du gène à la protéine.
CONCLUSION
IMPORTANCE DE LA MITOSE.
- – permettre le passage de l’état unicellulaire (zygote) à l’état pluricellulaire (embryon) ;
- – permettre ensuite le développement de l’embryon et la croissance du jeune individu ;
- – assurer le renouvellement des cellules qui meurent chez l’adulte.
- – Assurer la pérennité de l’espèce.
IMPORTANCE DE LA CELLULE.
La cellule est l’unité de structure et de fonctionnement de tout être vivant. Tous les êtres vivants, animaux et végétaux, sont constitués de cellules et chaque cellule provient d’une cellule : c’est la théorie cellulaire.
